Introduction
Le genre Dasypeltis est notamment connu pour son régime alimentaire ainsi que son mimétisme batésien avec les genres Echis et Causus. Les observations démontrent un schémas complexe entre l’aspect mimétique et défensif mis en oeuvre par le genre Dasypeltis en situation de danger.
Mots clés: comportement défensif, mimétisme, prédation, Dasypeltis, reptiles, organes vitaux
Présentation
Le genre Dasypeltis fait partie de la famille des colubrinaes. Son nom est dérivé du grec « dasy » (dense) et « péltê » (petit bouclier, par extension: plaque) et fait référence à une particularité de l’écaillure décrite par Wagler en 1830 (Ksas, Rémi, 2015). Il existe actuellement 13 espèces reconnues et 4 sous-espèces dispersées entre le complexe D. scabra et D. medici. Ce sont de petits serpents Africains inoffensifs dont la longueur totale varie entre 60cm et 100 cm. Ils sont exclusivement oophages. Leurs tête est ronde avec un cou non apparent. La pupille est verticalement elliptique. Ils ne possèdent aucune dent mais des vertèbres portants des hypapophyses qui leurs servent à percer la coquille des oeufs. La queue est courte. La coloration a une place importante dans l’identification des espèces qui sont morphologiquement très similaires (Chippaux Jean-Philippe, 2006).
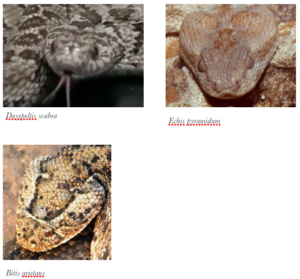
Dasypeltis est surtout connu pour son mimétisme batésien avec le genre Echis, bien que ce ne soit pas le seul genre. Cette forme de mimétisme est une évolution dans laquelle une espèce inoffensive imite les signaux d’alertes d’une espèce potentiellement nuisible à l’égard d’un ou plusieurs prédateurs communs (Bates, 1862 ; Aubret, Fabien et Mangin Alain, 2014). Le mimétisme batésien peut être visuel et/ou acoustique (Bates, 1862 ; Aubret, Fabien et Mangin Alain, 2014).
Tout comme d’autres vipéridaes et colubridaes, ils ont la capacité d’effectuer un signal d’avertissement par l’effet d’une respiration quadriphasique. La différence sonore entre les vipères et les serpents vipérines n’est pas distinguable (Aubret, Fabien et Mangin Alain, 2014). Cette dernière consiste à émettre un sifflement lors de l’expiration, suivi d’une pause puis d’un sifflement lors de l’inhalation, suivi à nouveau d’une pause de respiration (AUBRET, Fabien et MANGIN, Alain, 2014). Leur comportement dissuasif similaire au genre Echis est un avantage dans la survie de l’espèce, et ce, malgré le fait que l’aspect rond de la tête soit reconnaissable par les prédateurs (DELL’AGLIO, Denise Dalbosco, TOMA, Tiago Shizen Pacheco, MUELBERT, Adriane Esquivel, et al., 2012).
Matériels et méthodes
L’analyse comportementale a été effectuée sur un macbook air 13’’ d’avant 2015 avec le système d’exploitation Mac OS X Sierra. Le logiciel de montage vidéo iMovie à été employé pour ralentir les séquences à analyser.
Les vidéos proviennent du réseau socio-médiatique YouTube et d’un Dasypeltis atra captif filmé avec un Huawei P9 et une GoPro Hero 6 black. Douze vidéos sont analysées. Dasypeltis scabra représente 91,67 % soit un total de 11 vidéos. Dasypeltis atra représente 1 seule vidéo (8,33%). Le stimulus à l’origine du comportement défensif est dans 100% des cas une intervention humaine. 2 Vidéos (16,66%), Dasypeltis atra et un Dasypeltis scabra sont des individus captifs. 4 vidéos (33,2%) sont filmées dans un environnement artificiel tandis que le reste des vidéos (66,8%) est filmé dans le biotope naturel.
Les observations se portent sur la position initiale de l’espèce acculée ainsi que les mouvements effectués en réponses à ceux du stimulus. Toutes les observations ont pu être confirmées après lecture de ressources référencées.
Observations
Tous les Dasypeltis observés ont adopté une posture en S face au stimulus négatif. Bien que ceci ne soit pas systématique, c’était la condition inhérente à l’analyse comportementale de cette présente observation. L’aplatissement de la tête laisse apparaître le cou, notamment lors de la phase d’expiration, rendant la morphologie crânienne triangulaire. Cette observation a également été faite et confirmée post-analyse en 1982 par WERNER, YEHUDA L., et FRANKENBERG, ELIEZER. Immédiatement après ce dernier, un premier angle forme un demi-cercle sur environ un tiers du corps avant le début du deuxième tiers également en demi-cercle dans le sens inverse puis le 3ème arc de cercle. La queue, lorsqu’elle est plus grande, forme un quart de cercle derrière le troisième.
La distance avec le stimulus semble influencer la forme des demi-cercles précités. Plus le stimulus est proche, et plus l’individu resserre son corps allant jusqu’à laisser une ouverture de la largeur de la tête entre les angles du deuxième et troisième demi-cercle (Séquence 1). Sans pour autant se mettre en boule comme le ferait le Python régius, l’espace se rétracte à l’approche du stimulus (Séquence 2).

Toutes les tentatives de dissuasions par morsures n’ont pas été précédées d’un avertissement tel que l’ouverture de la gueule comme le ferait d’autres espèces, mais toutes ont prise fin en un très court laps de temps où la tête était cachée (Séquence 2) ainsi qu’un temps plus long elle ne faisait plus face au stimulus comme le montre les séquences 3 et 4 ci-dessous :

À contrario, toutes les tentatives d’intimidations par l’ouverture de la gueule n’ont pas été suivies d’attaque.
Durant la phase d’expiration, la morphologie triangulaire de la tête de D. scabra a été grandement accentuée présentant une similitude aussi proche de certaines vipères du genre Bitis et Causus que du genre Echis. Ceci peut en effet s’expliquer par la nécessité de se confondre le plus possible à une espèce nuisible car la tête triangulaire présente un nombre d’attaque inférieur aux têtes arrondies (DELL’AGLIO, Denise Dalbosco, TOMA, Tiago Shizen Pacheco, MUELBERT, Adriane Esquivel, et al., 2012).
La pupille semble, toujours durant la phase d’expiration, subir une contraction parfois verticale, parfois réductrice. Aucun élément visuel ou étudié ici ne permet de dire si ceci est un cas isolé, effectué volontairement par l’individu afin d’accentuer le mimétisme ou au contraire le résultat d’un schémas plus complexe lié à d’autres paramètres, par exemple, environnementaux.

Similitudes batésiennes connues
Une autre observation faite durant l’analyse des vidéos est la position à partir de laquelle l’individu s’élance pour « mordre ». L’ondulation émise par l’espèce afin d’émettre un son provoqué par le frottement des écailles les une contre les autres semble également influencé le moment ou l’individu se lancera en avant. Cette première phase d’observation a été effectuée sur la queue de l’animal au moment ou celle-ci commence à revenir vers le centre arrière de l’arc de cercle du troisième tiers de l’animal.
Discussions
Le comportement défensif des Dasypeltis est complexe et des études plus professionnelles semblent indispensables quant à une meilleure compréhension des changements physiologiques et physionomistes produits lorsqu’un individu se sent acculé.
Remerciements
Les auteurs référencés ci-dessous pour leurs études qui ont permis de confirmer et/ou d’étendre mes observations personnelles.
Sébastien Dubat, pour son engagement, sa disponibilité, ses nombreuses corrections et surtout sa méthodologie de travail qui me force continuellement à perfectionner la mienne.
Références
KSAS Rémi – L’étymologie des noms latins des serpents, VenomWorld Edition, 2015, p. 102.
CHIPPAUX Jean-Philippe – Les serpents d’Afrique occidentale et centrale. IRD éditions, 2006, p.113-114.
LOPEZ T. J., MAXSON L. R., et DOWLING H. G. – Phylogenetic relationships of the African egg-eating snake Dasypeltis scabra. Amphibia-Reptilia, 1993, vol. 14, no 3, p. 223-236.
DELL’AGLIO, DALBOSCO Denise, TOMA, Tiago SHIZEN PACHECO, MUELBERT, ESQUIVEL Adriane, et al. Head triangulation as anti-predatory mechanism in snakes. Biota Neotropica, 2012, vol. 12, no 3, p. 315-318.
MIMICRY, PROBABLE BEHAVIOURAL REINFORCEMENT OF BATESIAN, WERNER, YEHUDA L., et FRANKENBERG, ELIEZER – Department of Zoology, The Hebrew University of Jerusalem, Jerusalem 91904 Israel. ISRAEL JOURNAL OF ZOOLOGY, 1982, vol. 31, p. 137-150.
BATES Henry Walter. XXXII – Contributions to an Insect Fauna of the Amazon Valley. Lepidoptera: Heliconidae. Transactions of the Linnean Society of London, 1862, vol. 23, no 3, p. 495-566.
AUBRET Fabien et MANGIN Alain – The snake hiss: potential acoustic mimicry in a viper–colubrid complex. Biological journal of the Linnean Society, 2014, vol. 113, no 4, p. 1107-1114.